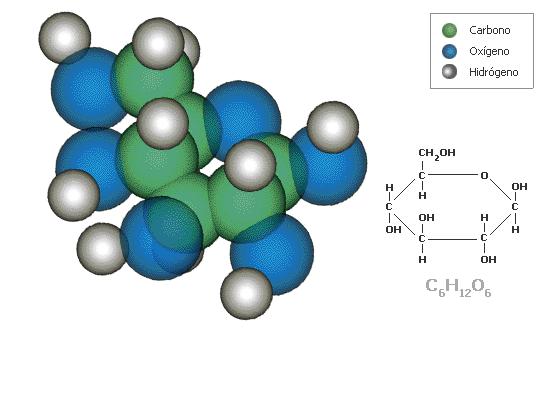
La glucólisis es un conjunto de reacciones que transforman la glucosa en piruvato. Es una vía casi universal. La glucólisis es una vía íntegramente citosólica.
Se distinguen 3 partes:
-Entrada de glucosa de todas las células. Sobretodo a músculos e hígado. Es el primer punto de control de la vía.
-Una vez ha entrado dentro del hepatocito:
Se puede estructurar en 2 etapas:
1. Pasar de Glucosa a Fructosa-1,6-difosfato.
2. Pasar de Fructosa-1,6-difosfato a dihidroxiacetona y gliceraldehido.
La primera fase es una fase de alteración química para dejar la molécula útil para la célula.
Primera fase de la glucólisis
1. La glucosa se fosforila en el alcohol por la hexoquinasa. La hexoquinasa une el fosfato del ATP mediante un enlace fosfoéster y la transforma en Glucosa-6-fosfato. La Glucosa-6-P está preparada para los procesos que continúan en la glucólisis. La fosforilación transforma 1 molécula neutra en una molécula cargada negativamente. Hace que ahora, la glucosa-6-P no pueda volver a salir por la membrana debido a su carga negativa.
2. A la célula no le gusta la forma aldosa y hace la forma cetosa (Fructosa-6-P). Es realizado por la fosfoglucosa isomerasa.
3. Implica la adquisición de un segundo fosfato que va a parar al C1 y forma la fructosa-1,6-bisfosfato. Lo relaiza la fosfofructoquinasa-1 (PFK-1). La fructosa-1,6-bisfosfato es completamente simétrica. La PFK-1 es un enzima clave en la glucólisis.
4. La fructosa-1,6-bisfosfato se parte por la mitad y da dos moléculas (dihidroxiacetona-fosfato(DHAP) y gliceraldehido-3-fosfato (G3P)). La glucólisis se da a partir del gliceraldehido-3-P. El equilibrio está desplazado hacia la dihidroxiacetona-fosfato. Sólo un 5% es Gliceraldehido-3-P. La desaparición contínua de G3P transforma la DHAP en G3P. Todo acaba siendo G3P.
Segunda fase de la glucólisis
A partir del G3P comienzan las transformaciones que dan lugar a Piruvato (Pyr) (reacciones de oxidación).
1. La primera reacción es que el enzima Gliceraldehido-3-Fosfatodeshidrogenasa oxida el grupo aldehido a ácido (gasta un NAD, que se reduce a NADH). Produce un enzima que es capaz de incorporar un fosfato al ácido carboxílico correspondiente. Se forma un éster. La fosforilación a nivel de sustrato se consigue porque hay ATP con un fosfato ya activado.
2. El 1,3-Bisfosfoglicerato cede 1 fosfato para sintetizar ATP. La reacción la cataliza la fosfogliceratoquinasa. El producto de la síntesis de ATP es el 3-fosfoglicerato. Sufre transformaciones que dan lugar a la síntesis de Pyruvato. El P3 debe pasar a la posición 2. Se consigue mediante el enzima fosfogliceromutasa. La mutasa tiene un fosfato activo, que se lo da a la posición 2. En un momento hay dos fosfatos, pero luego, se lo quita de la posición 3. Todas las mutasas funcionan así. Se hace para liberar el OH en la posición 3.
3. Se produce la deshidratación del OH. Se transforma el alcohol en enol, por la enolasa. Es parecido a Pyruvato, pero con enol y un fosfato. Se llama fosfoenolpiruvato (PEP). Es una molécula muy inestable que se transforma en Pyruvato por la Pyruvatoquinasa y se acopla la energía que se desprende para sintetizar ATP.
ANÁLISIS ESTEQUIOMÉTRICO DE LA GLUCÓLISIS
Glucosa + 2Pi + 2 ADP + 2 NAD+ 2 Pyr + 2 ATP + 2 NADH + 2 H+ + 2 H2O
La glucólisi es una vía que transforma la glucosa en Pyruvato y, a su vez, reduce 2 NAD+ del citosol a NADH y usa 2 Adp para formar 2 ATP.
La célula en cuanto puede, transforma otros monosacáridos a moléculas que están en la vía de la glucólisi.
Fructosa
Se encuentra en el azúcar, se metaboliza según:
· En el hígado: la fructosa se transforma en fructosa-1-P. Se gasta 1 ATP que pasa a ADP. Lo realiza la fructoquinasa. La fructosa-1-P es degradada por una aldolasa, que da lugar a dihidroxiacetona-P y a gliceraldehido. El gliceraldehido es fosforilado con gasto de ATP a G3P. Se produce el mismo rendimiento que en la glucosa.
· En el tejido adiposo: la fructosa es fosforilada a fructosa-6-P a partir de la transformación de ATP a ADP por la hexoquinasa. La hexoquinasa tiene mucha menos afinidad por la fructosa que por la glucosa. Por eso, en el hígado, la hexoquinasa fosforila glucosa. En el tejido adiposo la hexoquinasa (HK) actúa sobre la fructosa porque no hay tanta glucosa.
Galactosa
La galactosa se fosforila en Gal-1-P y se gasta ATP que pasa a ADP. Lo hace la galactoquinasa. La Gal-1-P es transferida al Uridindifosfoglucosa (UDPglucosa), que es la forma activada de la glucosa, por la Gal-1-P-uridiltransferasa y se genera UDPgalactosa y se libera Glucosa-1-P Después se transforma la Glucosa-1-P en Glucosa-6-P mediante la fosfoglucomutasa.
Para que el circuito funcione correctamente, hay que regenerar UDPglucosa. Hay una epimerasa que retransforma UDP-galactosa en UDPglucosa.
La galactosa rinde igual que la glucosa.
 La gluconeogénesis es una ruta metabólica anabólica que permite la síntesis de glucosa a partir de precursores no glucídicos. Incluye la utilización de varios aminoácidos, lactato, piruvato, glicerol y cualquiera de los intermediarios del ciclo de los ácidos tricarboxílicos (o ciclo de Krebs) como fuentes de carbono para la vía metabólica. Todos los aminoácidos, excepto la leucina y la lisina, pueden suministrar carbono para la síntesis de glucosa. Los Ácidos grasos de cadena par no proporcionan carbonos para la síntesis de glucosa, pues el resultado de su β-oxidación (Acetil-CoA) no es un sustrato gluconeogénico; mientras que los ácidos grasos de cadena impar proporcionarán un esqueleto de Carbonos que derivarán en Acetil-CoA y Succinil-CoA (que sí es un sustrato gluconeogénico por ser un intermediario del ciclo de Krebs).
La gluconeogénesis es una ruta metabólica anabólica que permite la síntesis de glucosa a partir de precursores no glucídicos. Incluye la utilización de varios aminoácidos, lactato, piruvato, glicerol y cualquiera de los intermediarios del ciclo de los ácidos tricarboxílicos (o ciclo de Krebs) como fuentes de carbono para la vía metabólica. Todos los aminoácidos, excepto la leucina y la lisina, pueden suministrar carbono para la síntesis de glucosa. Los Ácidos grasos de cadena par no proporcionan carbonos para la síntesis de glucosa, pues el resultado de su β-oxidación (Acetil-CoA) no es un sustrato gluconeogénico; mientras que los ácidos grasos de cadena impar proporcionarán un esqueleto de Carbonos que derivarán en Acetil-CoA y Succinil-CoA (que sí es un sustrato gluconeogénico por ser un intermediario del ciclo de Krebs).